| |
РЕКОМЕНДАЦИИ ПО ИСПОЛЬЗОВАНИЮ ГЕНЕТИЧЕСКИХ РЕСУРСОВ (БИОРАЗНООБРАЗИЯ) ПШЕНИЦ ДЛЯ СОЗДАНИЯ СОРТОВ НОВОГО ПОКОЛЕНИЯ С ИЗМЕНЕННОЙ АРХИТЕКТОНИКОЙ КОЛОСА.
В настоящее время очевидно, что не включенный в генофонд возделываемых видов пул генетической изменчивости (генов) как выведенных из возделывания видов сельскохозяйственных растений, так и родственных им видов, может быть использован для дальнейшего улучшения наиболее продуктивных современных сортов, в том числе основной продовольственной культуры для 30% населения Земли, мягкой пшеницы [Li A., Liu D., Yang W., Kishii M., Mao L Synthetic Hexaploid Wheat: Yesterday, Today, and Tomorrow // Engineering. 2018. Vol. 4. P. 552-558]. Считается, что такая стратегия селекции, основанная на широком использовании ранее невостребованных признаков и свойств, особенно связанных с изменением архитектоники вегетативных и генеративных частей широко возделываемых растений, а также генов, контролирующих их выраженность будет способствовать дальнейшему прогрессу селекции и развитию высокопродуктивного сельскохозяйственного производства [Гончаров Н.П., Кондратенко Е.Я., Вавилова В.Ю. Генетика адаптивности и архитектоника пшениц // Тез. всерос. конф. «Механизмы устойчивости растений и микроорганизмов к неблагоприятным условиям среды». Иркутск, 2018. С. 13-16]. Такой подход создания высоко адаптивных, стрессоустойчивых сортов нового поколения за счет использования более широкого генетического разнообразия и диверсификации сортов - особенно привлекателен для обеспечения продовольственной безопасности страны. Для его научного обеспечения необходимо получение новых знаний. Особенно это касается архитектоники растений будущего и создания модели сортов нового поколения [Гончаров Н.П., Гончаров П.Л. Скороспелость, диверсификация сортов и селекция будущего // Селекция с.-х. растений в аридных территориях Сибири и Дальнего Востока. Н-ск, 2015. С. 74-82]. При этом, структура колоса – один из важнейших признаков злаков, связанный с такими хозяйственно ценными их качествами, как продуктивность [Guo Z., Zhao Yu., Röde M.S. et al. Manipulation and prediction of spike morphology traits for the improvement of grain yield in wheat // Scientific Reports. 2018. Vol. 8. P. 14435. doi:10.1038/s41598-018-31977-3]. Колосья разных видов пшениц отличаются по форме, размерам, плотности, остистости, окраске и т.д. В последнее время отечественными селекционерами введены в культуру сорта видов пшениц, столетие уже как выведенные из возделывания Triticum sphaerococcum Perc., T. spelta L. и др. (рис.1).
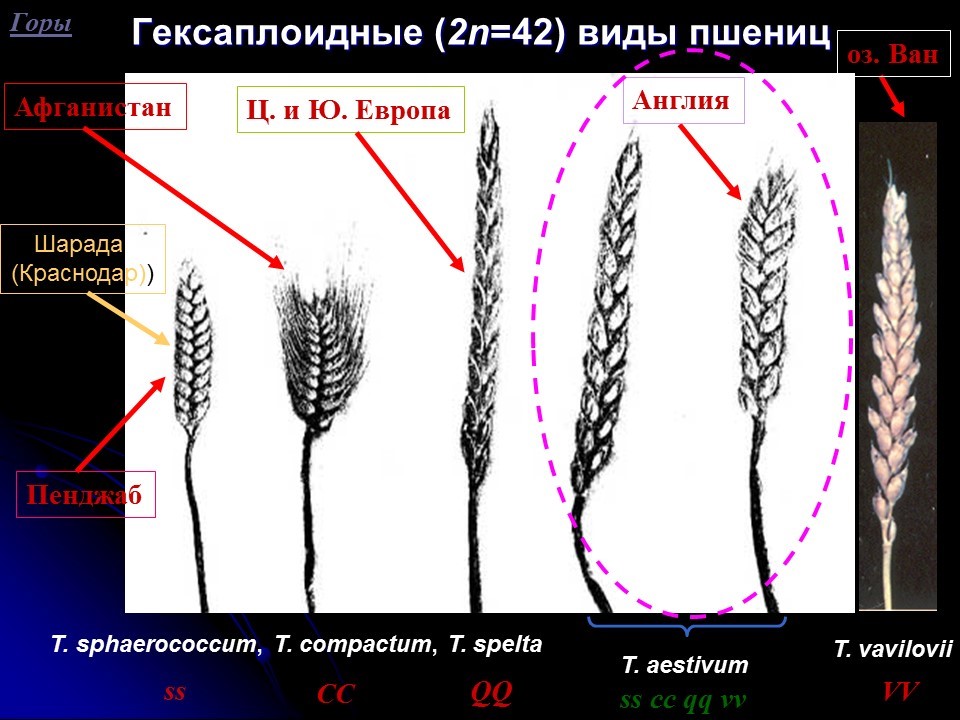
Рис.1. Гексаплоидные виды пшениц и гены контролирующие их таксономически важные признаки, определяющие архитектонику колоса
Известно, что процесс одомашнивания кардинально изменил архитектонику и свойства диких видов [Dubcovsky J., Dvořák J. Genome plasticity a key factor in the success of polyploid wheat under domestication // Science. 2007. Vol. 316. P. 1862–1866]. Признаки голозерность, отсутствие ломкоколосости и яровость сыграли ключевую роль в процессе доместикации и обеспечили успешное культивирование пшеницы на огромных пространствах [Dubcovsky J., Dvořák J. Genome plasticity a key factor in the success of polyploid wheat under domestication // Science. 2007. Vol. 316. P. 1862–1866].
Архитектоника растений непосредственно связана с урожайностью [Martinek P., Bednar J. Changes of spike morphology (multirow spike – MRS, long glumes – LG) in wheat (Triticum aestivum L.) and their importance for breeding // The Proceed. Intern. Conf. «Genetic Collections, isogenic and alloplasmic lines». Novosibirsk. 2001. P. 192–194]. У пшениц число генов, обусловливающих отличия в структуре колоса (его архитектоники), ограничено [Konopatskaia I., Vavilova V., Blinov A., Goncharov N.P. Spike Morphology Genes in Wheat Species (Triticum L.) // Proc. Latv. Acad. Sci. Sect. B Nat. Exact, Appl. Sci. 2016. V. 70. № 6. P. 345–355]. Большинство из них используется не только в селекции культуры, но и таксономии. При этом признаки, определяющие форму колоса у пшениц, относятся к таксономически значимым и обуславливают отличие одного таксона от другого. К числу основных/ключевых генов, контролирующих такие признаки, относят Q, C, S [Goncharov, 2011]. В настоящее время интрогрессия генов, обусловливающих отличия по архитектонике колоса из видов-сородичей (ячмень, рожь). К сожалению, доноры геномов В и D полиплоидных пшениц Aegilops speltoides Tausch и Ae. squarrosa L. (syn. Ae. tauschii Coss.) в этом плане практически не изучены. Один из возможных путей использование информации по архитектонике у хорошо изученных видов злаков, широко возделываемых в Старом Свете, таких как ячмень, рис и модельных объектов и поиск генов-гомологов (ортологов) у пшениц и/или перенос таковых молекулярно-биологическими методами.
Новая сборка последовательности генома мягкой пшеницы [Appels R., Eversole K., Feuillet C., et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome // Science. 2018. V. 361. № 6403. P.eaar7191] и одного из доноров ее элементарных геномов [Jia J., Zhao S., Kong X., et al. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation // Nature. 2013. V. 496. № 7443. P. 91–95. Marino R., Volante A., Brandolini A., Heun M. A high-resolution einkorn (Triticum monococcum L.) linkage map involving wild, domesticated and feral einkorn genotypes // Plant Breed. / ed. Abbo S. 2018. V. 137. № 5. P. 682–690] открывает перспективу для целенаправленного поиска и исследования новых хозяйственно-значимых признаков с помощью современных технологий редактирования генома, таких как CRISPR/Cas9, уже были ранее успешно опробованных на пшенице [Gil-Humanes J., Wang Y., Liang Z., et al. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR/Cas9 // Plant J. 2017. V. 89. № 6. P. 1251–1262. Liang Z., Chen K., Li T., et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes // Nat. Commun. 2017. V. 8. P. 14261]. Возможность создавать целенаправленные мутации в конкретных генах позволяет использовать информацию о гомологии генов с другими видами злаков для получения желаемых признаков без проведения «слепого» мутационного скрининга. Консервативные гены с высокой гомологией с большой вероятностью будут контролировать развитие сходных признаков, что позволяет предсказывать признак при внесении сходных мутаций в гомологичные гены. Например, у риса фенотип плотной и вертикально стоящей метелки (DEP, описано выше), вызванный мутацией в гене DEP1, проявляется за счет делеции участка в 637 пар нуклеотидов в середине пятого экзона [Huang X., Qian Q., Liu Z., et al. Natural variation at the DEP1 locus enhances grain yield in rice // Nat. Genet. 2009. V. 41. № 4. P. 494–497]. Исследование гомолога DEP1 на пшенице с нормальными или компактными колосьями показали, что изученные сорта обладают интактной последовательностью DEP1 [Vavilova V., Konopatskaia I., Kuznetsova A.E., et al. DEP1 gene in wheat species with normal, compactoid and compact spikes // BMC Genet. 2017. V. 18. № Suppl 1.], в тоже время другие гены, ответственные за компактный фенотип у пшеницы еще не были клонированы и молекулярно охарактеризованы. С помощью технологии CRISPR/Cas9 было бы возможно воспроизвести хозяйственно-значимый фенотип DEP риса у пшеницы, произведя аналогичную делецию в пятом экзоне гомолога DEP1.
Оценка характеристик колоса выполняется экспертом на основании визуального анализа колоса и в настоящий момент требует существенных затрат времени. Эффективность фенотипирования колосьев можно повысить за счет внедрения компьютерных технологий, организации хранения информации в базах данных, использования алгоритмов машинного обучения для анализа полученной информации [Генаев М.А., Комышев Е.Г., Фу Хао и др. SpikeDroidDB -информационная система для аннотации морфометрических характеристик колоса пшеницы // Вавиловский журнал генетики и селекции. 2018. Т. 22. № 1. С. 132-140.].
Составлены по результатам выполнения проекта РНФ №16-16-10021 руководителем проекта Н.П. Гончаровым и ответственным исполнителем К.В. Устьянцевым.
|
|
